눈과 망막
thumb|right|450px|
UNIQb4b28ac70c6647c5-nowiki-00000001-QINU1UNIQb4b28ac70c6647c5-nowiki-00000002-QINU
눈은 다른 감각들에 비해, 우리를 둘러싼 세상에 대해 비교할 수 없는 수준의 지각을 가능케 해주는 시각의 기관입니다. 눈은 주변 환경으로부터 광자(photon)을 수집하고 광자를 전기적 자극으로 바꿔주며 이렇게 바뀐 자극은 시각적으로 인지되는 영상으로 바뀌게 됩니다. 시각적 처리의 일부는 눈 안에서 행해지지만 대부분은 두뇌의 시각 피질(visual cortex)에서 이뤄지게 됩니다.
Contents
눈의 해부학적 구조(The Anatomy of the Eye)
각막(The Cornea)
각막은 동공과 홍채를 덮고 있는 눈의 투명한 표면입니다. 각막은 단순히 눈을 보호하는 기능뿐만 아니라 빛이 망막으로 가기 위해 지나가는 첫 번째 굴절 표면입니다. 건강한 각막은 혈관이 없으며 따라서 건강한 상태를 유지하기 위해 공기 중으로부터 산소를 받아들입니다. 각막에 산소공급이 오랜 기간 중단되면(가령 콘택트 렌즈를 허용 기간 이상 착용하는 경우), 인체는 각막에 혈관을 자라게 해 이러한 산소 부족을 보충하려 합니다. 이는 부정적인 면역 시스템 반응의 확률을 높이게 됩니다(충혈, 통증, 부종, 결국에는 각막의 영구 손상 등을 유발).
각막은 여섯 겹으로 이루어져 있으며 각각 상피세포, 보우만층, 기질, 두아층, 데스메막, 그리고 각막 내피세포층입니다(The epithelium, bowman's layer, stroma, dua's layer, Descemet membrane and corneal endothelium). [2]
상피세포층은 나머지 각막을 보호하는 역할을 하며 보우만 층 역시 마찬가지 입니다.[3], 기질층은 각막 두께의 90% 가량을 차지하며 투명합니다. 두아 층은 각막의 층들 중 가장 최근에 발견된 층으로 2013년도에 개별적인 층일 가능성이 보고 되었으며,[4] 아직 그 기능은 잘 알려지지 않았습니다. 데스메막과 내피세포 층은 각막과 눈의 나머지 부분들 간의 체액과 영영분 흐름을 조절하는 기능을 합니다.[5][6]
홍채와 동공(The Iris and the Pupil)
파일:300px-Eye_dilate.gif 홍채는 각막과 렌즈 사이에 위치한 눈의 일부분 입니다. 홍채의 중심에는 빛이 눈으로 들어올 수 있게 해주는 조리개인 동공이 있습니다. 홍채의 근육은 밝은 빛에 노출될 때 동공을 축소시키며 어두운 곳에서는 동공을 확대시키는 역할을 합니다. 멜라닌 색소의 정도가 홍채의 색깔을 결정하는 가장 중요한 요소 중 하나입니다. 멜라닌이 상대적으로 적은 경우, 홍채는 푸르거나 초록빛을 띠며 멜라닌이 많은 경우 홍채는 갈색 또는 검은색을 띄게 됩니다.[7]
렌즈(The Lens)
렌즈는 홍채 뒤에 있는 구조물로 빛을 모아 망막에 맺히게 합니다. 성인의 렌즈는 혈관이 없으며 수양액(aqueous humor)로부터 영양분을 받습니다. 그러나 발생단계의 자라나는 렌즈는, 보통 태어나기 전에 퇴행하는, 유리체 동맥(hyaloid artery)으로부터 영양을 공급받습니다. 모양체근이 렌즈의 모양을 바꾸어 빛이 렌즈를 통과해 망막에 맺힐 수 있도록 해주며 이러한 과정을 통해 눈이 다양한 거리에 있는 물체에 초점을 맞힐 수 있게 됩니다. [8]
The Vitreous Humour
The vitreous humour is a thick, gel-like fluid that maintains the shape of the eye, by filling the space between the lens and the retina and optic disk. It makes up about 80% of the volume of the eye, and is composed of about 98% water.[9]
Sclera
The sclera is more commonly known as the white of the eye. It is a white fibrous layer that becomes transparent at anterior part of the eye and forms the cornea. The human eye is repetitively rare in that the Iris is small enough that its position can be clearly seen against the sclera, allowing someone to tell where another individual is looking, which has developed into a form of non-verbal communication.
Anatomy of the human retina

The retina is a light-sensitive layer of tissue that lines the rear surface of the eye. Light from one's visual field passes through the eye and projects onto the retina to create an image. Subsequently, retinal neurons detect this image, which initiates a cascade of biochemical and electrical processing that is sent through the optic nerve and eventually to the visual cortex of the brain. These biochemical and electrical signals provide the basis for vision.
Cells of the Retina
Photoreceptors
- Cone cell en.png
The anatomy of a cone cell
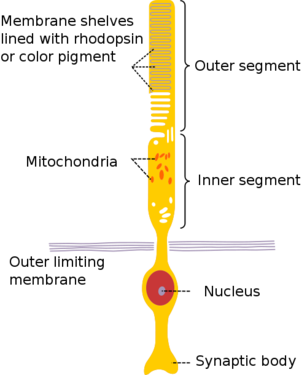
The anatomy of a rod cell[11]

Photoreceptors consist of two broad classes of cells: rods and cones. Rods are concentrated at the outer edges of the retina and are used in peripheral vision. They are more sensitive to light than cones, and are almost entirely responsible for night vision (also called scotopic vision). Cones are more concentrated towards the center of the retina, and are the only photoreceptor type found in the center of the retina (the fovea). Cones are responsible for color vision (also called photopic vision). Mammals usually have either two or three different types of cone cells, because in order to specify the wavelength of a stimulus (i.e., its color), the outputs of at least two cone types must be compared. These photoreceptor cells contain proteins called opsins in their outer segments (really just modified cillia) which bind a chromophore derived from Vitamin A, allowing these cells to convert absorbed light into an electrical impulse. Different wavelengths of light can be detected by varying the structure of the opsin protein expressed in the cell. [12]
Horizontal Cell

Horizontal cells are thought to exist in two types, each with a distinct shape, which together provide feedback to all photoreceptor cells. Despite the number of cells with which they form synapses, horizontal cells represent a relatively small population of the retina’s cells (less than 5% of cells of the inner nuclear layer). The specific reason for the existence of the two classes of horizontal cells is not yet known; it potentially involves detection of color differences in the red-green system.
Amacrine Cell

Amacrine cells appear to allow for ganglion cells to send temporally correlated signals to the brain: input to two separate ganglion cells from the same amacrine cell will tend to make those ganglion cells send signals at the same time. The amacrine cells whose behaviors are well understood have been shown to have very specific functions.
Bipolar Cell

Bipolar cells connect photoreceptors and ganglion cells. Their function is to transmit signals from photoreceptors to ganglion cells, either directly or indirectly. Bipolar cells get their name from their shape — they have a central cell body from which two different sets of neurites ( axons or dendrites) extend. They can make connections with either rods or cones (but not both simultaneously), and they also form connections with horizontal cells. Unlike most neurons, which communicate with one another using action potentials, bipolar cells “talk” with other cells using graded potentials.
Ganglion Cell

Ganglion cells are the output cells of the retina. Their axons leave the eye and travel through the optic nerve to the brain, sending the processed visual stimulus to the lateral geniculate nucleus, forming synapses onto neurons that project to the primary visual cortex, where the stimulus can be further interpreted.
Wiring it all together

{kind=link}
{kind=link}
Understanding the functions of the individual cells is the first step to understanding vision, but it does not begin to explain the complex processes that occur in our eyes providing us with the sense. Today, scientists are studying retinal connectomics to better understand vision. But, mapping the full connectome is too difficult a process for the technology currently available, instead, scientists map partial connectomes.
By looking at the connections between the neurons, scientists can understand how an input to one neuron can affect one that's connected to it. Scientists look at connections between multiple neurons, treating them as a system, and try to understand how an input affects the output. While scientists study systems of just a few neurons at a time, there is a basic understanding of how the retina processes information as a whole.[10]
A simplification of the process
To explain the function of the retina in the most basic way, four of the major types of retinal neurons will be considered, the photoreceptors, bipolar cells, amacrine cells, and ganglion cells. This wiki page will continue to expand the model to include more factors from the retinal neurons.
The first step to processing visual information is the collection of photons by the photoreceptors. The two types of photoreceptors, rods and cones, while serving different purposes, are similar in function.
In general, the rods and cones form synapses with the bipolar cells. While the cones can often be the only cone to form a synapse with a bipolar cell, many rods tend to form synapses with the same bipolar cell. Because multiple rods share a single bipolar cell, the sharpness of the image is traded off for sensitivity of the signal.
The bipolar cells then form synapses with amacrine cells. The amacrine cells provide signal feedback and transmit the signal to the ganglion cells.
The ganglion cells are the final step of the process in the processing of light in the retina. It is with the ganglion cells that the signals from the cones are compared to process what color something is. The ganglion cells transfer the signal out of the retina to the brain.[10]
Horizontal Cell Feedback
While horizontal cells represent a small portion of the retina, they do form many synapses with the photoreceptors. One of the leading theories on the function of horizontal cells is to increase the contrast between the light and dark regions. The horizontal cells will inhibit the signal of an excited cone and the cones surrounding it. While this dampens the signal, it also eliminates the signal from surrounding cones that aren't excited, so only the ganglion cells associated with the excited cone will be excited, rather than all the ganglion cells of the cone and its neighbors.
The rods receive feedback from specialized horizontal cells, which also give feedback to cones, in such a way that the two feedback systems are kept separate.[10]
Amacrine Cell Feedback
While most cells in the retina are flexible in their function, amacrine cells are highly specialized to do specific tasks.
Scientists have proposed that amacrine cells make ganglion cells fire in correlation with one another, which could increase the amount of information that can be transferred by the optic nerve.[10]
Current Research
According to many neuroscience textbooks, retinal ganglion cells can be categorized into two different types according to a property that is known as their receptive field. Neurons with receptive fields have been found in the auditory (hearing) system, the somatosensory (feeling) system and the visual system, and the receptive field of a particular neuron can generally be defined as a region of space in which the presence of a stimulus will alter the firing of that neuron.
In the visual system, a receptive field of a particular retinal ganglion cell is defined as the region of the photoreceptor cell layer in the retina that alters the firing (signal-sending) of that ganglion cell when it is stimulated with light. According to textbook accounts, retinal ganglion cells either have ON-center, OFF-surround or OFF-center, ON-surround receptive field. An ON-center, OFF-surround ganglion cell will send a signal when the center of its receptive field detects light, but will be inhibited from firing when the area surrounding the center (the surround) of its receptive field detects light; OFF-center, ON-surround cells have the exact opposite response to light stimulation.
{kind=link}
Within the last decade, it has become increasingly clear that the notion that only two types of receptive fields exist in photoreceptors is a gross oversimplification. Scientists now know that ganglion cells come in at least 15 or 20 types, each of which has a distinct shape and physiological function, and which correspondingly has connections with different types of cells in the rest of the retina.
At the Max Planck Institute (MPI) for Medical Research in Heidelberg, Germany, a dataset was obtained from a mouse retina in order to investigate this diversity in retinal ganglion cells – by applying two imaging techniques one after the other (two-photon microscopy (2P) and serial block-face scanning electron microscopy (SBFSEM)), scientists have been able to obtain images that show both neural activity and connectivity in retinal ganglion cells. However, the images are very difficult to analyze and interpret, and doing so is a very time-consuming process. Computer scientists at MIT are working on developing software to help with retinal image analysis, but computational analysis is currently much less accurate and reliable than that performed by humans.
The ultimate goal motivating the research on the retina that is being done at places like MPI and MIT is to use 2P and SBFSEM images in order to identify specific cell types within the broad classes of retinal cells that were described earlier, and further to understand connectivity between these cells. Only once the different cell types have been comprehensively catalogued will researchers be able to investigate their specific functions.
This is where YOU come in!
In order to fully understand retinal computation, it is necessary to map all of the connections that converge onto ganglion cells, as this diversity of connections generates the diversity of visual signals that are sent to the brain. The challenge now is to refine the coarse knowledge about retinal connectivity in order to gain a much more in-depth understanding of the specific functions of each and every cell type in the retina. Currently, scientists think that there are at least between fifty and sixty types, so there is much work to be done!
References
- ↑ Eye-diagram no circles border http://commons.wikimedia.org/wiki/File:Eye-diagram_no_circles_border.svg
- ↑ Binder, P. S. et al. (July 1991) High-voltage electron microscopy of normal human cornea. Invest. Ophthalmol. Vis. Sci. 32 (8): 2234-43
- ↑ Dua, Harminder S. et al. (September 2013) Human Corneal Anatomy Redefined: A Novel Pre-Descemet's Layer (Dua's Layer) Ophthalmology 120 (9): 1778-1785 doi: 10.1016/j.ophtha.2013.01.018
- ↑ McKee, Hamish D. et al. (May 2014, published online February 2014) Re: Dua et al.: Human corneal anatomy redefined: a novel pre-Descemet layer (Dua's layer) (Ophthalmology 2013;120:1778–85) Ophthalmology 121 (5): e24-e24 doi: 10.1016/j.ophtha.2013.12.021 \"Abstract: We read the recent claim of the discovery of a new corneal layer by Dua et al with incredulity.1 The existence of pre-Descemet stromal tissue remaining after pneumodissection is well known. Their further investigation of this pre-Descemet stroma confirms that it is stroma, and not a new corneal layer.\"
- ↑ McKee, Hamish D. et al. ANZ Cornea Meeting 2014 Abstracts, page 3 \"Dua’s layer” is just previously described pre-Descemet stroma ... Medical eponyms have traditionally been created by one’s peers to commemorate the importance of a person’s contribution and findings. Dua has taken an interesting step of creating his own eponym, even before his claims have stood the test of further investigation and scrutiny, and despite current trends to avoid medical eponyms (and when they are used, to use the nonpossessive form). If one prefers a medical eponym to describe the pre-Descemet stroma that remains after pneumodissection, then ‘the Feizi stroma’ would be more appropriate.\"
- ↑ Steinert, Roger (Medscape, 01 October 2014) A Controversy in Cornea \"... the purported discovery of a new layer in the cornea ... We all know that today it is quite frowned upon to use names of scientists to describe tissues. It's much more appropriate and helpful to use anatomic terms or physical terms that make sense.\"
- ↑ \"iris.\" Encyclopaedia Britannica. Encyclopaedia Britannica Online Academic Edition. Encyclopædia Britannica Inc., 2014. Web. 16 Jun. 2014. http://www.britannica.com/EBchecked/topic/294031/iris
- ↑ \"lens.\" Encyclopaedia Britannica. Encyclopaedia Britannica Online Academic Edition. Encyclopædia Britannica Inc., 2014. Web. 16 Jun. 2014. http://www.britannica.com/EBchecked/topic/336040/lens
- ↑ "human eye." Encyclopaedia Britannica. Encyclopaedia Britannica Online Academic Edition. Encyclopædia Britannica Inc., 2014. Web. 16 Jun. 2014. http://www.britannica.com/EBchecked/topic/1688997/human-eye/64878/The-transparent-media?anchor=ref531532
- ↑ 10.0 10.1 10.2 10.3 10.4 Masland, R. The fundamental plan of the retina (2001). Nature Neuroscience 4 (9): 877-886
- ↑ Human Physiology and Mechanisms of Disease by Arthur C. Guyton (1992) p.373
- ↑ Plachetzki, D.; Fong, C.; Oakley, T. (2010). "The evolution of phototransduction from an ancestral cyclic nucleotide gated pathway". Proceedings. Biological sciences / the Royal Society 277 (1690): 1963–1969. doi:10.1098/rspb.2009.1797. PMC 2880087. PMID 20219739.
- ↑ Kolb, Helga, Nelson, Ralph, Fernandez, Eduardo, Jones, Bryan, The Organization of the Retina and Visual System, Simple Anatomy of the Retina. http://webvision.med.utah.edu/book/part-i-foundations/simple-anatomy-of-the-retina/
- ↑ http://en.wikipedia.org/wiki/Receptive_field
{kind=link}