El ojo y su retina

- cuerpo vítreo
- ora serrata
- músculo ciliar
- zónula ciliar
- canal de Schlemm
- pupila
- cámara anterior
- córnea
- iris
- corteza del cristalino
- núcleo del cristalino
- proceso ciliar
- conjuntiva
- músculo oblicuo inferior
- músculo recto inferior
- músculo recto medio
- arterias y venas retinianas
- papila óptica
- duramadre
- arteria central de la retina
- vena central de la retina
- nervio óptico
- vena vorticosa
- vaina bulbar
- mácula
- fóvea
- esclerótica
- coroides
- músculo recto superior
- retina
El ojo es el órgano de la vista, que nos permite percibir nuestro entorno en una escala superior comparada con los demás sentidos. Los ojos captan los fotones del ambiente y los convierten en impulsos eléctricos, posteriormente integrados en imágenes visuales. Parte del proceso ocurre en el ojo, pero la mayoría de las etapas tienen lugar en la corteza visual, en el encéfalo.
Contents
Anatomía del ojo
La córnea
La córnea es la superficie traslúcida del ojo que recubre a la pupila y el iris. Además de brindar protección, es la primera superficie de refracción por la que la luz llega a la retina. La córnea carece de irrigación sanguínea en condiciones normales, de modo que el oxígeno necesario lo obtiene del aire. Cabe resaltar que ante la ausencia prolongada de oxígeno, como en el empleo excesivo de lentes de contacto, el organismo trata de compensarla con angiogénesis corneal. Esto influye en la posible aparición de una respuesta inmunológica perjudicial (enrojecimiento, dolor, hinchazón y, al final, lesión corneal irremediable).
La córnea tiene cinco capas: epitelio, membrana de Bowman, estroma, membrana de Descemet y endotelio. El epitelio y la membrana de Bowman protegen los demás componentes de la córnea. El estroma representa 90 % del grosor total en tanto sea transparente. La membrana de Descemet y el endotelio regular el flujo de líquido y nutrientes entre la córnea y el resto del ojo. [2]
En 2013 se reportó la posible existencia de una sexta capa, con la denominación propuesta de membrana de Dua,[3], aunque algunos investigadores han tomado esta aseveración «con escepticismo» y alegan que no es una capa nueva, sino parte del estroma,[4] y también han debatido en nombre propuesto[5][6]
El iris y la pupila
El iris es la zona del ojo entre la córnea y el cristalino. En el centro se encuentra la pupila, una apertura que permite la entrada de luz. Los músculos del iris contraen el iris ante luz brillante y lo dilatan en condiciones de poca luz. La concentración de melanina es el factor preponderante que define el color del iris. En la ausencia relativa de melanina, el color del iris suele ser azul o verde. Por otra parte, con mayor cantidad de melanina, es más probable que el iris sea café o negro.[7]
The Lens
The lens is a structure behind the iris that focuses light onto the retina. The adult lens contains no blood vessels and is nourished by the aqueous humour, but during development the growing lens has nutrients supplied by the hyaloid artery, which usually regresses before birth. The ciliary muscles change the shape of the lens to focus light which passes through onto the retina allowing the eye to focus on objects that are at varying distances.[8]
The Vitreous Humour
The vitreous humour is a thick, gel-like fluid that maintains the shape of the eye, by filling the space between the lens and the retina and optic disk. It makes up about 80% of the volume of the eye, and is composed of about 98% water.[9]
Sclera
The sclera is more commonly known as the white of the eye. It is a white fibrous layer that becomes transparent at anterior part of the eye and forms the cornea. The human eye is repetitively rare in that the Iris is small enough that its position can be clearly seen against the sclera, allowing someone to tell where another individual is looking, which has developed into a form of non-verbal communication.
Anatomy of the human retina

The retina is a light-sensitive layer of tissue that lines the rear surface of the eye. Light from one's visual field passes through the eye and projects onto the retina to create an image. Subsequently, retinal neurons detect this image, which initiates a cascade of biochemical and electrical processing that is sent through the optic nerve and eventually to the visual cortex of the brain. These biochemical and electrical signals provide the basis for vision.
Cells of the Retina
Photoreceptors
- Cone cell en.png
The anatomy of a cone cell
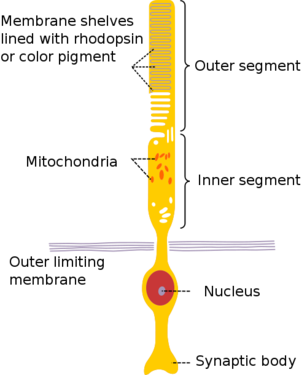
The anatomy of a rod cell[11]

Photoreceptors consist of two broad classes of cells: rods and cones. Rods are concentrated at the outer edges of the retina and are used in peripheral vision. They are more sensitive to light than cones, and are almost entirely responsible for night vision (also called scotopic vision). Cones are more concentrated towards the center of the retina, and are the only photoreceptor type found in the center of the retina (the fovea). Cones are responsible for color vision (also called photopic vision). Mammals usually have either two or three different types of cone cells, because in order to specify the wavelength of a stimulus (i.e., its color), the outputs of at least two cone types must be compared. These photoreceptor cells contain proteins called opsins in their outer segments (really just modified cillia) which bind a chromophore derived from Vitamin A, allowing these cells to convert absorbed light into an electrical impulse. Different wavelengths of light can be detected by varying the structure of the opsin protein expressed in the cell. [12]
Horizontal Cell

Horizontal cells are thought to exist in two types, each with a distinct shape, which together provide feedback to all photoreceptor cells. Despite the number of cells with which they form synapses, horizontal cells represent a relatively small population of the retina’s cells (less than 5% of cells of the inner nuclear layer). The specific reason for the existence of the two classes of horizontal cells is not yet known; it potentially involves detection of color differences in the red-green system.
Amacrine Cell

Amacrine cells appear to allow for ganglion cells to send temporally correlated signals to the brain: input to two separate ganglion cells from the same amacrine cell will tend to make those ganglion cells send signals at the same time. The amacrine cells whose behaviors are well understood have been shown to have very specific functions.
Bipolar Cell

Bipolar cells connect photoreceptors and ganglion cells. Their function is to transmit signals from photoreceptors to ganglion cells, either directly or indirectly. Bipolar cells get their name from their shape — they have a central cell body from which two different sets of neurites ( axons or dendrites) extend. They can make connections with either rods or cones (but not both simultaneously), and they also form connections with horizontal cells. Unlike most neurons, which communicate with one another using action potentials, bipolar cells “talk” with other cells using graded potentials.
Ganglion Cell

Ganglion cells are the output cells of the retina. Their axons leave the eye and travel through the optic nerve to the brain, sending the processed visual stimulus to the lateral geniculate nucleus, forming synapses onto neurons that project to the primary visual cortex, where the stimulus can be further interpreted.
Wiring it all together

{kind=link}
{kind=link}
Understanding the functions of the individual cells is the first step to understanding vision, but it does not begin to explain the complex processes that occur in our eyes providing us with the sense. Today, scientists are studying retinal connectomics to better understand vision. But, mapping the full connectome is too difficult a process for the technology currently available, instead, scientists map partial connectomes.
By looking at the connections between the neurons, scientists can understand how an input to one neuron can affect one that's connected to it. Scientists look at connections between multiple neurons, treating them as a system, and try to understand how an input affects the output. While scientists study systems of just a few neurons at a time, there is a basic understanding of how the retina processes information as a whole.[10]
A simplification of the process
To explain the function of the retina in the most basic way, four of the major types of retinal neurons will be considered, the photoreceptors, bipolar cells, amacrine cells, and ganglion cells. This wiki page will continue to expand the model to include more factors from the retinal neurons.
The first step to processing visual information is the collection of photons by the photoreceptors. The two types of photoreceptors, rods and cones, while serving different purposes, are similar in function.
In general, the rods and cones form synapses with the bipolar cells. While the cones can often be the only cone to form a synapse with a bipolar cell, many rods tend to form synapses with the same bipolar cell. Because multiple rods share a single bipolar cell, the sharpness of the image is traded off for sensitivity of the signal.
The bipolar cells then form synapses with amacrine cells. The amacrine cells provide signal feedback and transmit the signal to the ganglion cells.
The ganglion cells are the final step of the process in the processing of light in the retina. It is with the ganglion cells that the signals from the cones are compared to process what color something is. The ganglion cells transfer the signal out of the retina to the brain.[10]
Horizontal Cell Feedback
While horizontal cells represent a small portion of the retina, they do form many synapses with the photoreceptors. One of the leading theories on the function of horizontal cells is to increase the contrast between the light and dark regions. The horizontal cells will inhibit the signal of an excited cone and the cones surrounding it. While this dampens the signal, it also eliminates the signal from surrounding cones that aren't excited, so only the ganglion cells associated with the excited cone will be excited, rather than all the ganglion cells of the cone and its neighbors.
The rods receive feedback from specialized horizontal cells, which also give feedback to cones, in such a way that the two feedback systems are kept separate.[10]
Amacrine Cell Feedback
While most cells in the retina are flexible in their function, amacrine cells are highly specialized to do specific tasks.
Scientists have proposed that amacrine cells make ganglion cells fire in correlation with one another, which could increase the amount of information that can be transferred by the optic nerve.[10]
Current Research
According to many neuroscience textbooks, retinal ganglion cells can be categorized into two different types according to a property that is known as their receptive field. Neurons with receptive fields have been found in the auditory (hearing) system, the somatosensory (feeling) system and the visual system, and the receptive field of a particular neuron can generally be defined as a region of space in which the presence of a stimulus will alter the firing of that neuron.
In the visual system, a receptive field of a particular retinal ganglion cell is defined as the region of the photoreceptor cell layer in the retina that alters the firing (signal-sending) of that ganglion cell when it is stimulated with light. According to textbook accounts, retinal ganglion cells either have ON-center, OFF-surround or OFF-center, ON-surround receptive field. An ON-center, OFF-surround ganglion cell will send a signal when the center of its receptive field detects light, but will be inhibited from firing when the area surrounding the center (the surround) of its receptive field detects light; OFF-center, ON-surround cells have the exact opposite response to light stimulation.
{kind=link}
Within the last decade, it has become increasingly clear that the notion that only two types of receptive fields exist in photoreceptors is a gross oversimplification. Scientists now know that ganglion cells come in at least 15 or 20 types, each of which has a distinct shape and physiological function, and which correspondingly has connections with different types of cells in the rest of the retina.
At the Max Planck Institute (MPI) for Medical Research in Heidelberg, Germany, a dataset was obtained from a mouse retina in order to investigate this diversity in retinal ganglion cells – by applying two imaging techniques one after the other (two-photon microscopy (2P) and serial block-face scanning electron microscopy (SBFSEM)), scientists have been able to obtain images that show both neural activity and connectivity in retinal ganglion cells. However, the images are very difficult to analyze and interpret, and doing so is a very time-consuming process. Computer scientists at MIT are working on developing software to help with retinal image analysis, but computational analysis is currently much less accurate and reliable than that performed by humans.
The ultimate goal motivating the research on the retina that is being done at places like MPI and MIT is to use 2P and SBFSEM images in order to identify specific cell types within the broad classes of retinal cells that were described earlier, and further to understand connectivity between these cells. Only once the different cell types have been comprehensively catalogued will researchers be able to investigate their specific functions.
This is where YOU come in!
In order to fully understand retinal computation, it is necessary to map all of the connections that converge onto ganglion cells, as this diversity of connections generates the diversity of visual signals that are sent to the brain. The challenge now is to refine the coarse knowledge about retinal connectivity in order to gain a much more in-depth understanding of the specific functions of each and every cell type in the retina. Currently, scientists think that there are at least between fifty and sixty types, so there is much work to be done!
References
- ↑ Diagrama del ojo commons.wikimedia.org/wiki/File:Eye-diagram_no_circles_border.svg
- ↑ Binder, P. S. et al. (July 1991) High-voltage electron microscopy of normal human cornea. Invest. Ophthalmol. Vis. Sci. 32 (8): 2234-43
- ↑ Dua, Harminder S. et al. (September 2013) Human Corneal Anatomy Redefined: A Novel Pre-Descemet's Layer (Dua's Layer) Ophthalmology 120 (9): 1778-1785 doi: 10.1016/j.ophtha.2013.01.018
- ↑ McKee, Hamish D. et al. (mayo de 2014, publicado en línea en febrero de 2014) Re: Dua et al.: Human corneal anatomy redefined: a novel pre-Descemet layer (Dua's layer) (Ophthalmology 2013;120:1778–85) Ophthalmology 121 (5): e24-e24 doi: 10.1016/j.ophtha.2013.12.021 «Resumen: Leímos con escepticismo la reciente declaración del descubrimiento de Dua y cols. de una nueva capa corneal. Se sabe que queda estroma residual antes de la capa de Descemet una vez hecha la neumodisección. Su investigación de este estroma pre-Descemet corrobora que es estroma, no una nueva capa corneal».
- ↑ McKee, Hamish D. et al. ANZ Cornea Meeting 2014 Abstracts, página 3 «La “capa de Dua” es el ya descrito estroma pre-Descemet […] los epónimos médicos se han creado tradicionalmente como reconocimiento de colegas a los aportes y hallazgos de alguien. Dua ha sacado ventaja creando su propio epónimo, antes de que siquiera sus declaraciones hayan pasado la prueba de la investigación minuciosa y el escrutinio, aunado a la reciente tendencia de evitar los epónimos médicos (y cuando ya están en uso, hacerlo en su forma no posesiva [en inglés]). Si se prefiere el epónimo del estroma remanente del estroma pre-Descemet una vez hecha la neumodisección, “estroma de Feizi” es lo más adecuado».
- ↑ Steinert, Roger (Medscape, 01 October 2014) A Controversy in Cornea «[…] el supuesto descubrimiento de una nueva capa en la córnea […] todos sabemos que actualmente se censura el uso de epónimos para nombrar tejidos. Es más adecuado y útil el empleo de términos anatómicos o físicos que sean acordes».
- ↑ "iris." Encyclopaedia Britannica. Encyclopaedia Britannica Online Academic Edition. Encyclopædia Britannica Inc., 2014. Web. 16 Jun. 2014.
- ↑ "lens." Encyclopaedia Britannica. Encyclopaedia Britannica Online Academic Edition. Encyclopædia Britannica Inc., 2014. Web. 16 Jun. 2014. http://www.britannica.com/EBchecked/topic/336040/lens
- ↑ "human eye." Encyclopaedia Britannica. Encyclopaedia Britannica Online Academic Edition. Encyclopædia Britannica Inc., 2014. Web. 16 Jun. 2014. http://www.britannica.com/EBchecked/topic/1688997/human-eye/64878/The-transparent-media?anchor=ref531532
- ↑ 10.0 10.1 10.2 10.3 10.4 Masland, R. The fundamental plan of the retina (2001). Nature Neuroscience 4 (9): 877-886
- ↑ Human Physiology and Mechanisms of Disease by Arthur C. Guyton (1992) p.373
- ↑ Plachetzki, D.; Fong, C.; Oakley, T. (2010). "The evolution of phototransduction from an ancestral cyclic nucleotide gated pathway". Proceedings. Biological sciences / the Royal Society 277 (1690): 1963–1969. doi:10.1098/rspb.2009.1797. PMC 2880087. PMID 20219739.
- ↑ Kolb, Helga, Nelson, Ralph, Fernandez, Eduardo, Jones, Bryan, The Organization of the Retina and Visual System, Simple Anatomy of the Retina. http://webvision.med.utah.edu/book/part-i-foundations/simple-anatomy-of-the-retina/
- ↑ http://en.wikipedia.org/wiki/Receptive_field