J-RGC

J-RGCs are retinal ganglion cells defined by their expression of the protein Junction Adhesion Molecule B (JAM-B). J-RGCs are direction selective OFF RGCs, responding specifically to upward motion in the visual field.[1] They can be recognized by their asymmetric dendritic arbors aligned in a dorsal-to-ventral direction across the retina. Like many direction selective ganglion cells, they project heavily to the superior colliculus (SC), but also innervate the dorsal lateral geniculate nucleus (dLGN). This was the first cell type in the retina to be classified based on the presence of a molecular biomarker (?check).
Contents
Anatomy
Shape

In about 85% of JAM-B cells, there exists a marked asymmetry in the dendritic arbor. In these cases, more than 90 percent of the dendritic arbor lies on one side of the soma. Asymmetry of the dendritic arbor is an unusual feature for a retinal ganglion cell because most of the dendrites of retinal ganglion cells are symmetrical. Therefore, the prevalence of this asymmetry in such a high proportion of JAM-B cells is notable. Even more notable is that the asymmetrical dendrites of JAM-B cells all point in the same direction, about 13 degrees nasal of ventral. Relative to the optic nerve head, there exists a longitudinal, rather than radial, dendritic orientation (dorsal to ventral).[1]
Alternatively, approximately 15% of JAM-B cells are not markedly asymmetrical. These non-asymmetrical JAM-B cells are almost all located near the dorsal and ventral margins of the retina. Therefore, nearly all JAM-B cells in the nasal, central, and temporal regions of the retina are markedly asymmetrical [1]
Connections

The dendrites of JAM-B cells have been found to arborize in a narrow band between processes of dopaminergic and cholinergic amacrines. Indeed, the dendrites of both symmetric and asymmetric JAM-B cells are confined to the outer third of the inner plexiform layer.
Retinal ganglion cells whose dendrites arborize in the outer half of the Inner Plexiform Layer are generally known to be OFF-RGCs--i.e., they respond to a decrease of light in the center of the receptive field.[3] Notably, several subcategories of OFF bipolar cell have processes in the same sublaminae as J-RGC dendrites, while processes of ON bipolar cells have been found to overlap minimally with J-RGCs. Thus the arborization pattern of J-RGCs led Kim et al. to first hypothesize that these cells are OFF retinal ganglion cells.[1]
J-RGC dendrites branch several times in the middle portion of the inner plexiform layer before reaching to the outer third of the inner plexiform layer. It is the outer third in which further branches and terminal arbors reside. Dendrites of both asymmetrical and symmetrical populations branch a few times in the middle of the IPL, then ascend to the outer margin, where further branches and terminal arbors reside in SL2 (see figure).[2]
Kim et al. used yellow fluorescent protein to trace the projection of JAM-B cells to the mouse brain. They found that JAM-B cells project heavily to the superior colliculus. Thus it seems that mice have invested heavily in the detection of upward motion. It remains to be determined why exactly this is the case.[1]
Physiology
Kim et al. found that J-RGCs exhibited three notable physiological asymmetries. These asymmetries, in order of their discussion here, are the direction of displacement of receptive field surround (and related asymmetric light response), the preferred direction for movement of a spot of light, and the space-time slant within the receptive field center.[1]
Light Response and its Correlation to Dendrite Orientation
JAM-B cells are OFF retinal ganglion cells, meaning that they respond to a decrease in light levels. Kim et al. probed J-RGCs with flashing lights and found that almost all fired when the light turned off. On the other hand, RGCs lacking evidence of JAM-B were nearly equally likely to be ON or OFF cells.
Furthermore, the receptive field of J-RGCs is unusual in that the integrated strength of its ON surround exceeds that of the OFF centre. Kim et al. reached this conclusion after recording the response of J-RGCs to different sizes of flashing light spots centered at the soma (cell body). When the flashing spot was of a certain large enough size, the J-RGC sometimes failed to respond. Upon increasing the size of flashing spot even further, Kim et al. found that the J-RGCs tended to respond at light onset. Thus the strength of the JAM-B cells' ON surround exceeds that of their OFF center--an unusual feature in a retinal ganglion cell.
Studies imply that no sizable populations of OFF RGCs exist with asymmetric dendrites pointing in directions other than dorsal-to-ventral.[2]
Upward Motion Selectivity
JAM-B cells respond selectively to upward motion. To test the direction sensitivity of each sample J-RGC, Kim et al. moved a small spot through its receptive field center along eight straight-line trajectories of different orientation. They found that the response correlated strongly with the direction of the spot's motion. Furthermore, the direction eliciting the most response matched the general direction in which the asymmetric dendritic tree pointed away from the J-RGC soma. In contrast, it had previously been found that direction-selective ganglion cells generally lack a correlation between physiological sensitivity and structural asymmetry.[1]
Kim et al. proposed that the asymmetry of a JAM-B cell's dendritic arbors is related to this direction selectivity, stating, "across the entire J-RGC population, the degree of direction selectivity was correlated with the degree of asymmetry of the receptive field. It has been found in another study that when probed with a small moving spot, the firing rate of J-RGCs correlated strongly with the direction of motion. The preferred direction of motion corresponds to the direction of the dendritic arbor from the soma"[2] As Kim et al. claim, "within a single molecularly defined class of OFF-RGCs, dendritic structure and cell function are closely linked, suggesting that the latter arises from the former." [1]
Whereas the selectivity of other direction-selective RGCs depends on input from starburst amacrine cells, which themselves show directional responses, J-RGCs receive little input from these cells and thus must rely on other mechanisms. One possible mechanism is suggested by the finding that inhibitory synapses on some RGCs are concentrated at distal dendrites. Distal inhibition on the asymmetric dendrites of J-RGCs could account for their asymmetrically displaced surround.[1]
Features of the JAM-B Receptive Field Shaping Direction Selectivity

The ON surround of a JAM-B cell is typically asymmetric, in that it is shifted in the direction of preferred motion relative to the center. The receptive field center was found by Kim et al. to possess an OFF portion with an additional ON "overshoot." This ON feature of the center combines functionally with the ON surround such that a light moving in the preferred direction across the receptive field will pass through the combined ON features of center and surround. If the light is moving at the right speed, the time delay of the center invokes a superposition of the excitation of the ON and OFF regions, thus eliciting an enhanced response (see figure). It is this mechanism which purportedly allows an OFF RGC to respond to bright lights with strong direction-selectivity.
Furthermore, Kim et al. found that the OFF center is biased in the space-time plane such that it is longer in the direction of preferred motion. This slanting of the OFF center renders the region more responsive to objects moving in the preferred direction (as opposed to objects moving perpendicular to the preferred direction.
An additional finding of Kim et al. is that the space-time slope of the OFF center is significantly greater than that of the ON-surround. As a result, a dark spot elicits a response of higher magnitude from the J-RGC when moving quickly, while a light spots elicits the greatest response at low speeds[1]
Molecules
JAM-B Cells are classified by the expression of the Junctional adhesion molecule B (JAM-B) protein.
Junctional adhesion molecule B is a protein that in humans is encoded by the JAM2 gene. JAM2 has also been designated as CD322 (cluster of differentiation 322). Tight junctions represent one mode of cell-to-cell adhesion in epithelial or endothelial cell sheets, forming continuous seals around cells and serving as a physical barrier to prevent solutes and water from passing freely through the paracellular space. The protein encoded by this immunoglobulin superfamily gene member is localized in the tight junctions between high endothelial cells. It acts as an adhesive ligand for interacting with a variety of immune cell types and may play a role in lymphocyte homing to secondary lymphoid organs [4].
Development
The pattern by which retinal ganglion cells develop in the retina varies by subtype. It has been found that J-RGCs develop via a gradual restriction of a diffuse pattern. The dendrites of J-RGC cells have been found to extend throughout the entire inner plexiform layer at P5 in mice. At P8 in mice it was found that branches had been pruned in the inner portion of the inner plexiform layer, while in the outer portion of the Inner Plexiform Layer the branches had propagated. In fact, they were found to have their arbors centered around starburst amacrines in SL3. Gradually the arbor distributions were found to shift outward through the IPL until P12, when they had achieved their adult pattern of restriction to LS2.[2]
- Error creating thumbnail: Unable to save thumbnail to destination
The development of retinal ganglion cells varies by cell type. Here we see J-, BD-, W3, and W7-RGCs at stages P5, P8, and P12/13. Note the J-RGC's gradual shifting of dendritic arbor distribution to the outer portion of the IPL.[2]
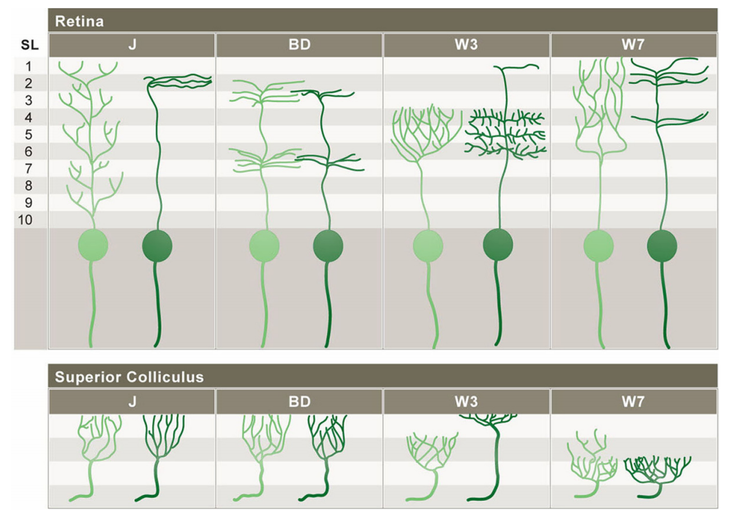
A visual overview of subtype-specific patterns of development in J-, BD-, W3-, and W7-RGCs. The light green figures represent RGC patterns at an immature age and the dark green figures represent the mature patterns. In addition to having specific patterns of dendritic lamination, retinal ganglion cell subtypes have distinctive axonal arbor patterns within the superior colliculus and lateral genticulate nucleus.[2]

History
JAM-B cells were first discovered by In-Jung Kim, Yifeng Zhang, Masahito Yamagata, Markus Meister, and Joshua R. Sanes of Harvard's Department of Molecular and Cellular Biology and Center for Brain Science.
To mark JAM-B cells for structural and functional study, they generated mice that expressed a ligand–activated Cre recombinase oestrogen receptor fusion protein 10 (CreER) under the control of regulatory elements from the JAM-B gene.
Before the discovery of JAM-B cells, retinal ganglion cells lacked a labeling system based on molecular markers. Thus Retinal Ganglion Cells were categorized by nonselective factors or else treated as a single population. Thus RGCs were often either subject to subjective classification or inaccurate overgeneralization.[1] Today, RGC subtypes in addition to JAM-B cells have been classified based on reporter genes. These subtypes include BD-, W3-, and W7-RGCs. These subsets can be identified using transgenic lines to mark them.
Open Questions/Status
Relationship between J-RGCs and direction-selective collicular cells
The collicular termination of J-RGCs (of which JAM-B cells are a majority) is intriguing in light of a study in which Drager and Hubel mapped the receptive fields of neurons in the superior colliculus of the mouse. Nearly all of the direction-selective neurons they studied (35 out of 38) preferred upward motion in the visual field. This preference corresponds to that of J-RGCs.[1] Kim et al. have proposed that the receptive fields of direction-selective collicular cells are built from J-RGCs. Research remains to be done on this topic.
Sensitivity to upward motion in mice
It is uncertain why the mouse seems to have invested so heavily in sensitivity to upward motion. Kim et al. have suggested mating JAM-B–CreER mice to other transgenics bearing appropriate Cre-activated channels or toxins. Thus it could be possible to inactivate the pathway and thereby directly test its function.[1]
References
- ↑ 1.00 1.01 1.02 1.03 1.04 1.05 1.06 1.07 1.08 1.09 1.10 1.11 1.12 1.13 1.14 1.15 In-Jung Kim et al. Molecular identification of a retinal cell type that responds to upward motion (2008). Nature 452: 478-482
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 2.6 2.7 In-Jung Kim et al. Laminar Restriction of Retinal Ganglion Cell Dendrites and Axons: Subtype-Specific Developmental Patterns Revealed with Transgenic Markers (2010). The Journal of Neuroscience. 30 (4): 1452-1462
- ↑ Wässle, H. Parallel processing in the mammalian retina (2004). Nat Rev Neuroscience. 5: 747-757
- ↑ JAM2
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||

{kind=link}